How Microbial Tryptamines MODULATE CONCIOUSNESS in Autism

By Christian Bogner, MD
Reading time ~40min
Listen to this article
Duration: 37min55sec
Disclaimer
This website does not provide medical advice. The information, including but not limited to, text, graphics, images, and other material, contained on this website is for educational purposes only. The purpose of this article is to promote broad consumer understanding and knowledge of various health topics. It is not intended to be a substitute for professional medical advice, diagnosis, or treatment. Always seek the advice of your physician or other qualified health care providers with any questions you may have regarding a medical condition or treatment before undertaking a new health care regimen, and never disregard professional medical advice or delay in seeking it because of something you have read on this website!
- Section 1: A Child who Cannot Speak, but can See Everything
- Section 2: The Gut is Not a Side Quest in Autism
- Section 3: Tryptophan, the Crossroads Amino acid
- Section 4: The Liver as a Psychedelic Conversion Hub
- Section 5: The Receptor Map of a Chronic Psychedelic State
- Section 6: Sensory Gating, Thalamus, and the Eye Contact Paradox
- QEEG Studies Reveal Striking Parallels
- Section 7: Speech Apraxia and Motor Planning, Autism’s Hidden Core
- Section 8: OCD Loops, Aggression, Sleep disruption, and Seizures, One Circuit Logic
- MRI and EEG patterns that fit an Altered Consciousness Model
- The Brain Overgrowth Phenomenon
- Section 9: Predictions, the Responder Profile, and Why this is Likely not a Niche Subgroup
- Section 10: Gut Balancing, what Alex built, what he Saw, and Why it Matters
- Section 11: Why we are Taking this to the Laboratory Now
- Resources
Section 1: A Child who Cannot Speak, but can See Everything
Three years ago I met Jacob, a 7 year old boy diagnosed with severe autism. He was non verbal, intensely hyperactive, and stimming nearly constantly. His behaviors were severe enough that most people around him assumed his internal world was sparse, or disorganized, or simply unreachable.
Then I met his incredible parents, Scott & Ashley and…the real Jacob.
After his school went virtual during Covid, Jacob learned to type at home with his patient mom. Once he had this reliable output channel figured out, he revealed a mind that was not only present, but astonishingly structured. He could track complex ideas, ask precise questions, and discuss chemistry with me at a level that forced me to stop and recalibrate what I thought I was looking at. He contributed to a clinical discovery that later became central in many families’ detoxification support conversations, sodium thiosulfate. Since then, hundreds of parents have reported meaningful improvements after carefully implementing sodium thiosulfate protocols as part of broader biomedical programs. This was just my first encounter with him.
Jacob also offered statements that, whether taken literally or understood as phenomenological descriptions, are profoundly instructive. He said he understands every language. He described having an IQ of 500+. He described meeting other individuals like him through telepathic communication. The point is not whether every claim can be externally verified. The point is that Jacob, among numerous others I have met, does not sound like a child with “no cognition.” He sounds like someone trapped behind a brokeninterface.
That single clinical pattern changes everything: what if autism, especially severe autism, is often not a disorder of intelligence, but a disorder of access?
If that is true, then the highest leverage question becomes brutally specific: what biological mechanism could preserve comprehension and internal complexity, while disrupting speech, motor planning, sensory gating, sleep, emotional regulation, and seizure threshold?
In the next section, the story moves away from the brain for a moment, because the most consequential chemistry in autism may begin in the gut.
Section 2: The Gut is Not a Side Quest in Autism
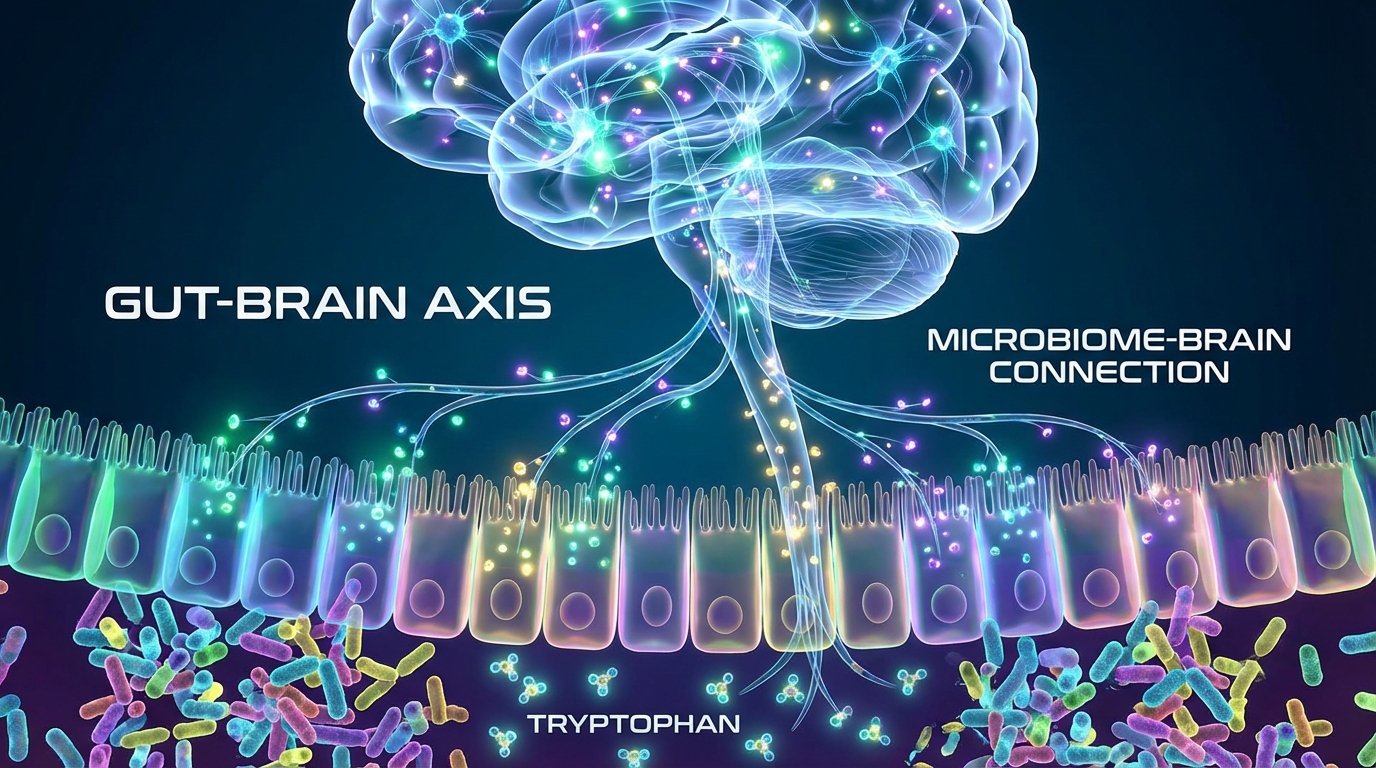
A large majority of individuals with autism have gastrointestinal dysfunction. Constipation, diarrhea, abdominal pain, reflux, food selectivity, and inflammatory patterns are not occasional comorbidities, they are often continuous, life shaping constraints.
For years, mainstream framing treated this as secondary. Brain first, gut second. Behavior first, biology later. That hierarchy is increasingly indefensible. The gut is not merely a digestive tube, it is a sensory organ, an immune organ, a neuroendocrine organ, and a biochemical factory that produces signaling molecules capable of reshaping the nervous system.
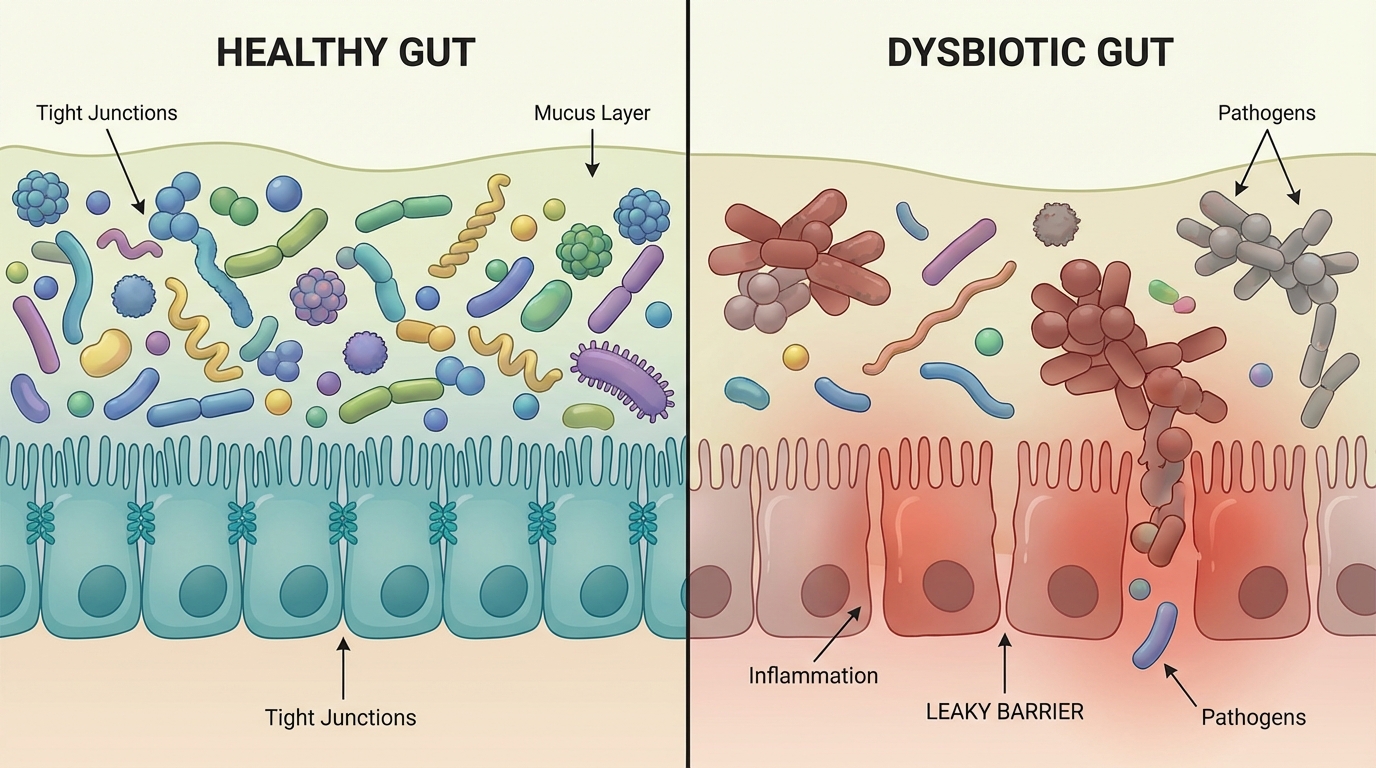
The microbiome, the community of bacteria and other microbes living in the gastrointestinal tract, behaves like a metabolic organ. It takes dietary inputs and host secretions, then outputs a chemical atmosphere. Some of that chemical atmosphere supports barrier integrity, immune tolerance, and normal neuromodulation. Some of it, under dysbiotic conditions, becomes inflammatory, neuroactive, and destabilizing.
If you accept one core premise, the rest becomes inevitable: when gut barrier function is compromised, the bloodstream becomes a delivery highway for molecules that were never supposed to reach systemic circulation at scale. Lipopolysaccharide (LPS), mycotoxins, metals, industrial toxins, oxalates, food protein are some examples. But in autism, there is another class of molecules that may be even more explanatory, because they do something that looks eerily like the autism phenotype we observe clinically.
They alter consciousness.
This is the foundation of the Endogenous Psychedelic Hypothesis, alternately described as microbiome driven psychedelic apraxia: A dysbiotic microbiome may actually be able to divert tryptophan metabolism toward the production of psychoactive indole tryptamines, which then chronically modulate brain circuits through the same receptor logic as classical psychedelics.
In the next section, the entire hypothesis turns on one biochemical crossroads, tryptophan.
Section 3: Tryptophan, the Crossroads Amino acid
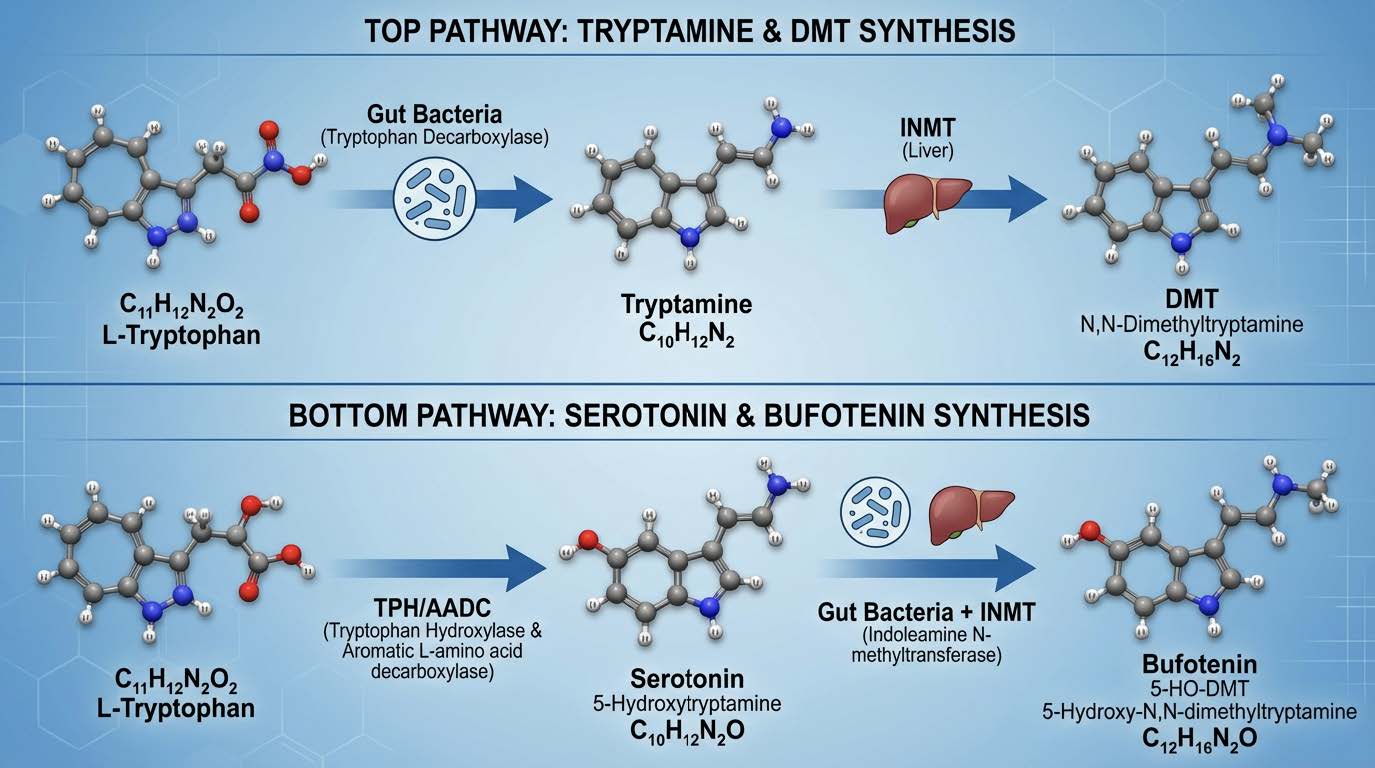
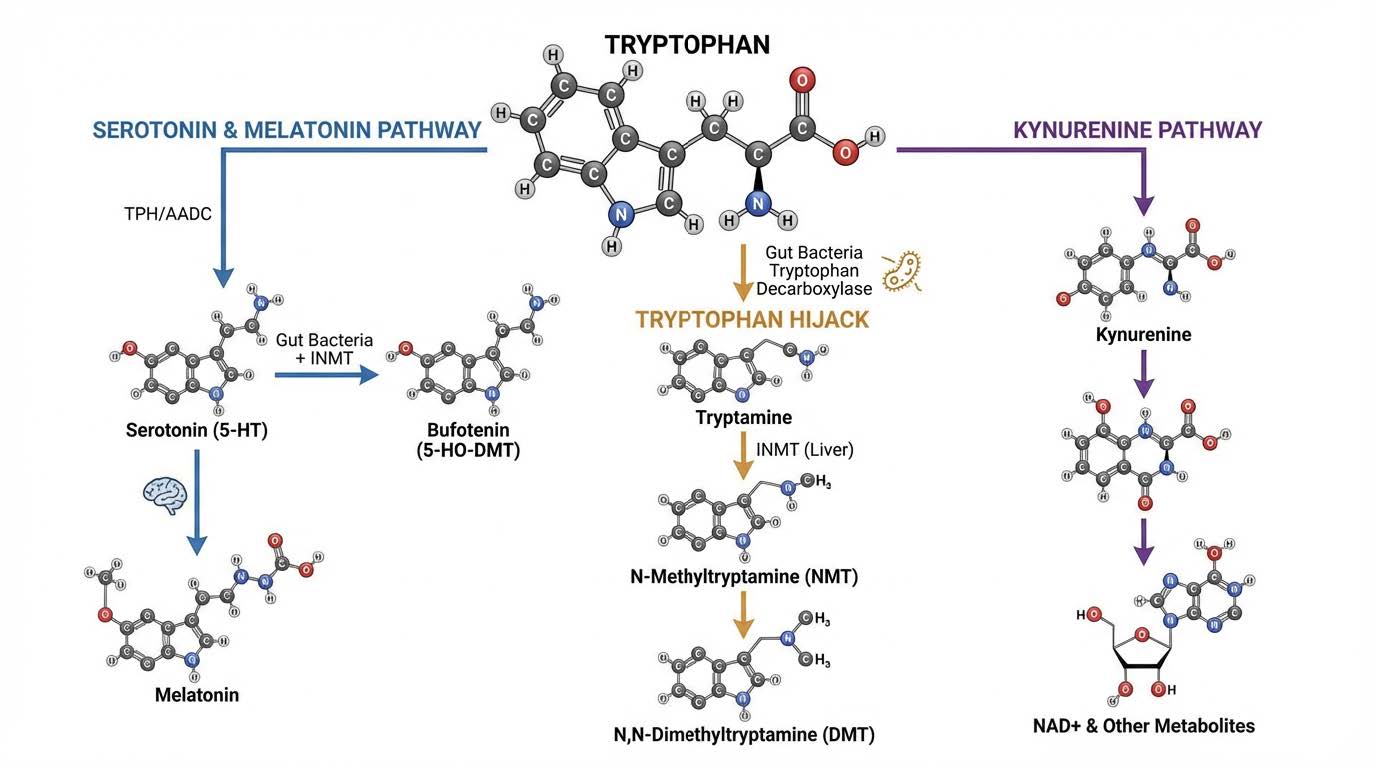
Tryptophan is not just a dietary amino acid. It is a strategic substrate. The body must decide where to send it. Under stable physiology, most tryptophan flows into the kynurenine pathway, contributing to NAD production, cellular energetics, and redox homeostasis. A smaller fraction goes toward serotonin production, e.g. helping with GI motility (GI produced serotonin does not cross into the brain). Another fraction can be transformed into diverse indole derivatives, some beneficial, some not.
In a dysbiotic gut, however, a different pathway can dominate: direct decarboxylation of tryptophan into tryptamine.
This is not speculative chemistry. Many bacterial species possess tryptophan decarboxylase activity. When those expand (we do see this clinically) and when competing protective species are depleted, the gut begins to behave like a tryptamine reactor.
A tryptophan hijack, orchestrated by bacteria. The microbiome diverts tryptophan away from the routes that support metabolic stability (consequence: mitochondria problems) and motility (consequence: IBS like symptoms), and instead generates an amine that is intrinsically neuroactive and, more importantly, can be converted into compounds with psychedelic receptor activity.
Once tryptamine production is high, three other realities amplify the risk.
- First, leaky gut increases absorption of these molecules.
- Second, liver enzymology can transform them into stronger compounds.
- Third, genetic polymorphisms can reduce breakdown and clearance.
This is where autism stops looking like a purely neurodevelopmental label and starts looking like a systems level metabolic state.
In the next section, we follow tryptamine out of the gut and into the liver, where it can be upgraded into DMT like chemistry.
Section 4: The Liver as a Psychedelic Conversion Hub
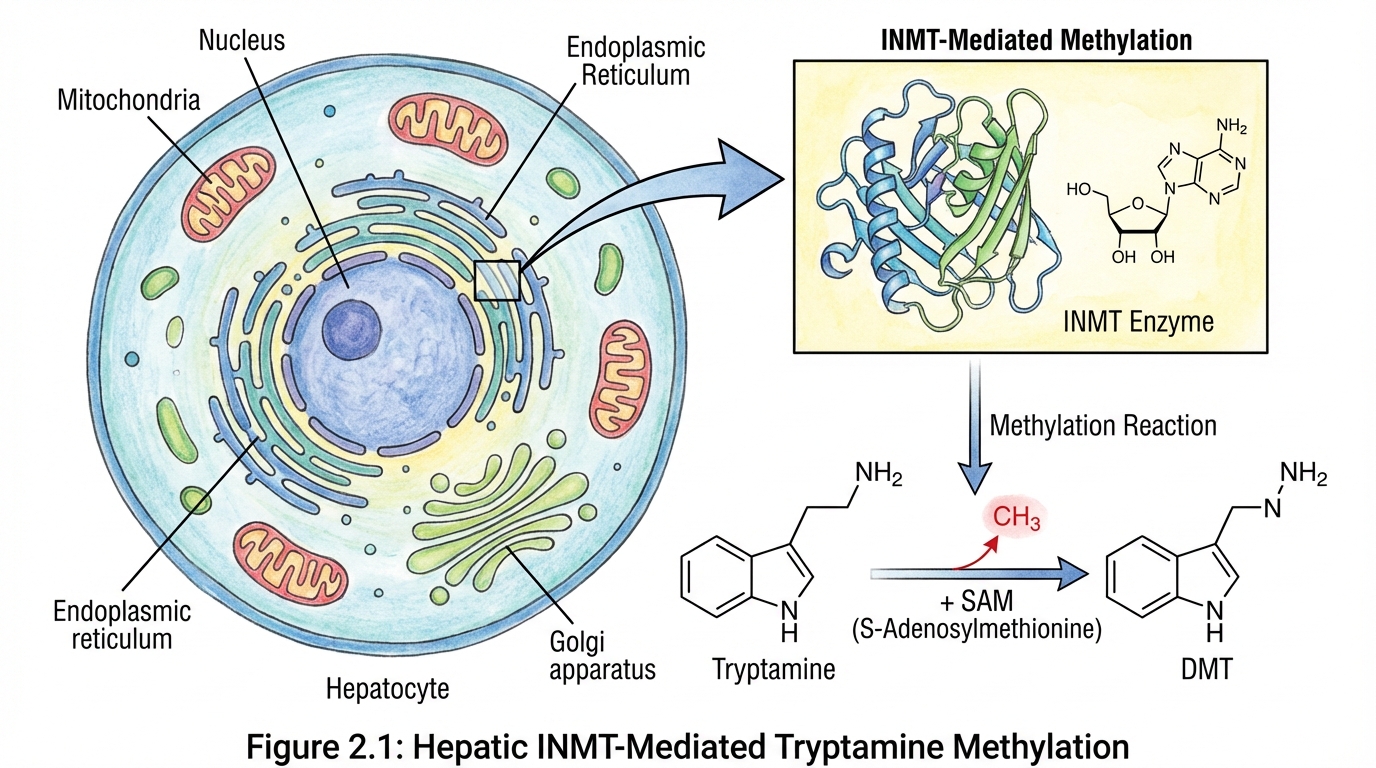
Tryptamine itself is biologically active. But the hypothesis becomes far more provocative when you follow what the body can do with tryptamine once it crosses into circulation.
The liver expresses high levels of an enzyme called indolethylamine-N-methyltransferase, commonly abbreviated INMT. INMT can methylate tryptamine into N-methyltryptamine and then into N,N dimethyltryptamine, DMT.
A parallel route also exists: serotonin can be methylated into 5 hydroxy DMT, commonly called bufotenin.
DMT and bufotenin are not obscure molecules. They sit at the center of psychedelic neuroscience. They are potent agonists of the serotonin 5HT2A receptor, and DMT also has meaningful activity at sigma 1 receptors, among other targets.
Now integrate the gut facts with the liver facts. If a dysbiotic microbiome drives tryptamine production, and if barrier function is compromised, and if INMT activity is present, then the body has the machinery to create psychedelic tryptamines endogenously.
Now let’s integrate genetics. Monoamine oxidase A, MAOA, is one of the critical enzymes involved in degradation of monoamines. If MAOA activity is lower due to polymorphisms, then clearance of these indole amines can be impaired. MAOA is X linked, which creates an elegant vulnerability structure: males have one X chromosome, which can magnify the impact of low activity variants, likely contributing to the male predominance in autism. Additional polymorphisms in methylation related genes can further destabilize handling of these metabolites and the broader neuroimmune terrain.
This section is where the hypothesis shifts from “interesting” to “mechanistically dangerous.” The body may be producing molecules that, in other contexts, are used explicitly to dissolve perception, alter salience, distort sensory gating, and disrupt motor sequencing.
In the next section, we translate that chemistry into receptor level neuroscience, because the receptor map explains why autism looks the way it does.
Section 5: The Receptor Map of a Chronic Psychedelic State

Classical psychedelics share a core mechanism: activation of the serotonin 5HT2A receptor. LSD, psilocin, mescaline, DMT, and many related compounds converge on that receptor logic, producing downstream effects that include cortical glutamate release, increased network flexibility, altered sensory processing, and a characteristic shift in consciousness.
If endogenous DMT and bufotenin are elevated chronically, the brain is not “hallucinating” in a cartoon sense. It is being forced into a different operating regime.
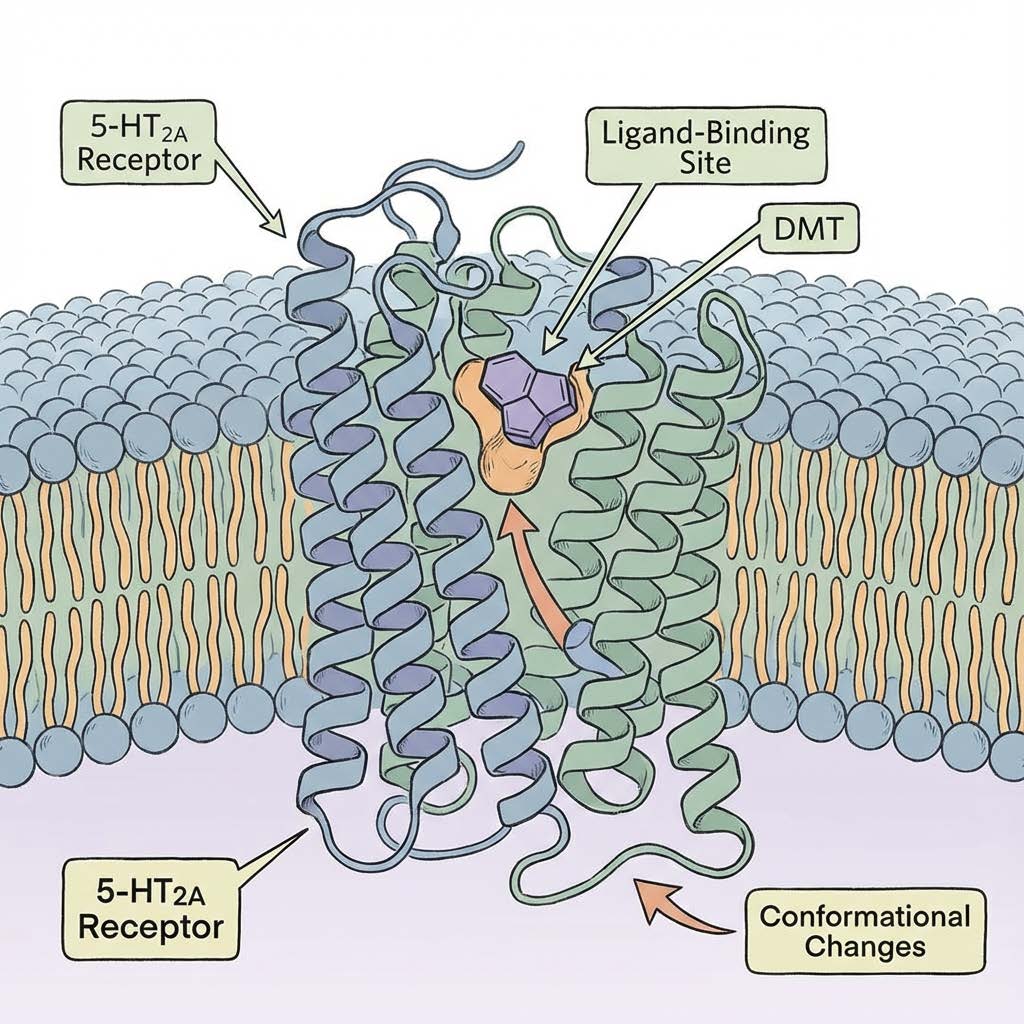
Here is what matters clinically. 5HT2A receptors are not sprinkled randomly. They are placed strategically in circuits that govern:
- Sensory gating, especially thalamocortical filtering
- Predictive coding and salience assignment
- Cortical excitation-inhibition balance
- Motor planning and sequencing through cortico-striato-thalamo-cortical loops
- Speech motor output through frontal motor planning areas and corticobulbar pathways
- Emotion regulation through limbic-cortical integration
- Sleep architecture through serotonergic regulation and melatonin synthesis competition
Sigma 1 receptors add another layer. Sigma 1 receptors sit at the endoplasmic reticulum mitochondrial interface and regulate calcium signaling, redox homeostasis, cellular stress responses, and synaptic plasticity. DMT is a sigma 1 agonist. If chronic endogenous DMT activity exists, sigma 1 overstimulation becomes a plausible contributor to altered connectivity, excitability, sensory processing, and resilience thresholds.
This is where a “behavioral diagnosis” starts looking like a specific neurochemical state.
Consider a striking clinical coincidence: the two FDA approved medications commonly used for autism irritability, risperidone and aripiprazole (Abilify), both block 5HT2A receptors. There are hundreds of receptor types in the human brain. The fact that US government approved pharmacology in autism leans heavily on 5HT2A antagonism is not proof, but it is directionally consistent with the idea that excessive 5HT2A signaling is involved.
In the next section, we go to the thalamus, because sensory gating and hypersensitivity are among the clearest “psychedelic like” features in autism, and they can explain eye contact avoidance in a new way.
Section 6: Sensory Gating, Thalamus, and the Eye Contact Paradox
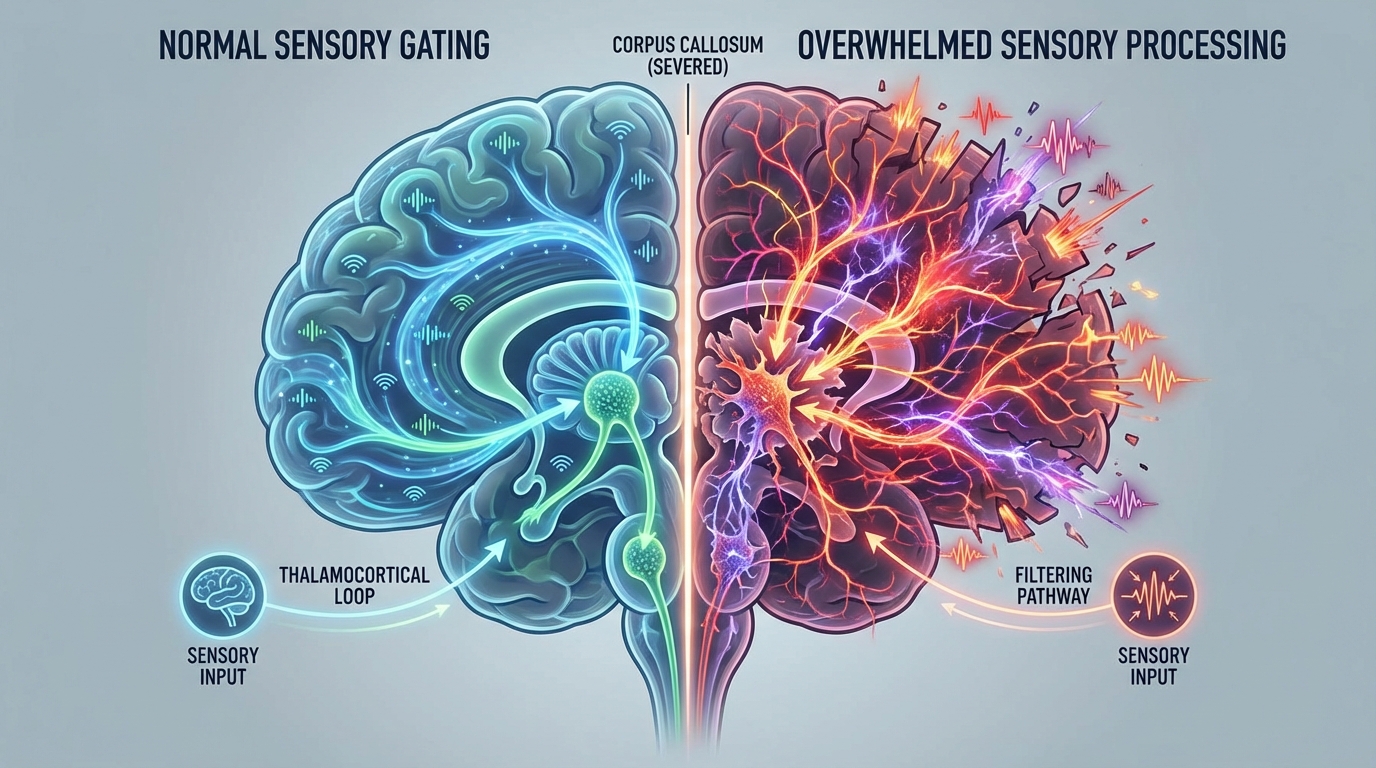
Many autistic individuals are not “uninterested” in the world. They are flooded by it. The clinical language, hypersensitivity, sensory overwhelm, startle reactivity, noise intolerance, tactile defensiveness, fits a nervous system that cannot filter.
The thalamus is one of the primary biological filters. It decides what gets amplified and what gets suppressed. The reticular thalamic nucleus modulates thalamocortical oscillations and contributes to attentional gating. Serotonin receptors, including 5HT2A and related subtypes, influence these dynamics.
QEEG Studies Reveal Striking Parallels
Quantitative EEG (QEEG) research provides objective support for the “chronic altered state” hypothesis. A key oscillatory signature of robust sensory gating involves alpha rhythms (8–12 Hz). In neurotypical brains, alpha waves often signify a state of “idling” or suppression of irrelevant sensory input.
- Reduced Alpha Suppression: Both psychedelic states (psilocybin, LSD, DMT) and autism are repeatedly associated with reduced alpha power and altered thalamocortical synchrony. This reduction suggests a failure of “top-down” inhibitory control, allowing excessive “bottom-up” sensory information to reach the cortex unchecked.
- Theta and Gamma Abnormalities: QEEG studies in autism frequently show increased theta power (4–8 Hz) in frontal and central regions, alongside altered gamma activity (>30 Hz). These patterns mirror findings in acute psychedelic states, where “entropic” or disorganized brain activity correlates with the dissolution of normal perceptual boundaries.
This convergent electrophysiological data strongly suggests that the autistic brain is operating in a regime of heightened excitability and reduced filtering—a functional mirror of the psychedelic state.
A key oscillatory signature here involves alpha rhythms, roughly 8 to 12 Hz, and broader alpha beta dynamics up to around 30 Hz, which are linked to sensory suppression and top down control. Psychedelic states are repeatedly associated with reduced alpha power and altered thalamocortical synchrony. Autism literature repeatedly reports abnormal oscillatory patterns, atypical sensory suppression, and altered thalamocortical connectivity.
Now connect phenomenology to circuitry. If a child lives in a chronic, low grade psychedelic state, then eye contact is not socially “confusing,” it is neurochemically overwhelming. Many psychedelic subjects describe direct eye contact as too intense, penetrating, emotionally exposing, hyper salient.
Reinterpret the classic autism feature: reduced eye contact may be a defensive strategy against excessive salience and sensory amplification, not a lack of social interest.
This same model also fits the pattern of sudden aggression, emotional volatility, and shutdown, e.g. when sensory gating fails, the nervous system is forced into primitive survival outputs.
In the next section, we move from perception to action, because the strongest proof point for this hypothesis is not only hypersensitivity. It is apraxia, motor planning failure, and the tragedy of speech trapped behind a broken motor interface.
Section 7: Speech Apraxia and Motor Planning, Autism’s Hidden Core
Autism is often described primarily as a social communication disorder. In practice, severe autism frequently looks like a motor disorder with downstream social consequences.
Motor impairments are extremely common. Dyspraxia, clumsiness, poor coordination, hypotonia, gait abnormalities, repetitive movements, oral motor dysfunction, and profound speech impairment are not rare add ons. They are often central.
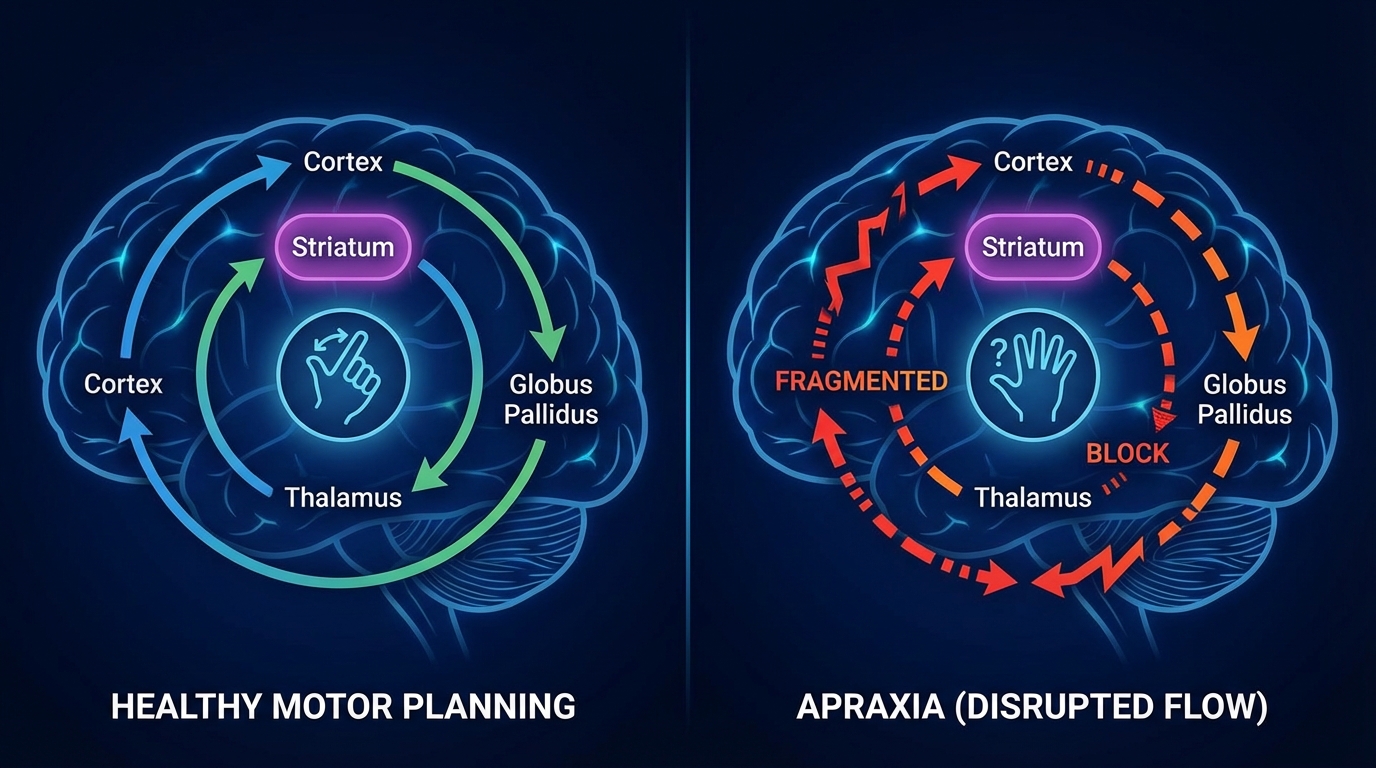
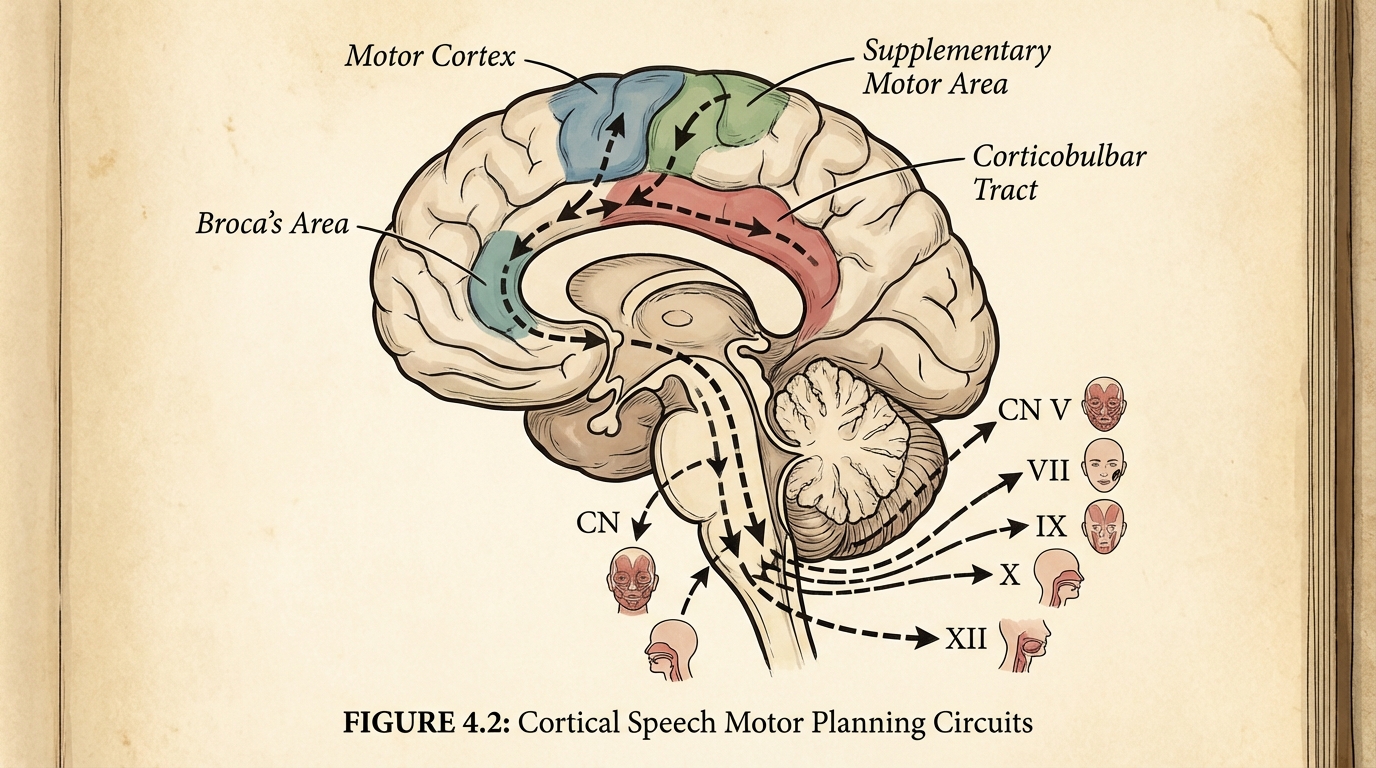
Motor planning is not “muscle strength.” Motor planning is circuit logic: the ability to sequence, initiate, and execute fine grained actions through cortico striato thalamo cortical loops, cerebellar error correction, and frontal motor planning networks.
Now insert psychedelic neuropharmacology. DMT and bufotenin modulate basal ganglia activity and thalamocortical gating. Psychedelics are well known to disrupt fine motor sequencing, reaction time, and coordinated output. Acute psychedelic states can produce ataxia, tremor, erratic movements, and speech disruption.
Speech apraxia in autism may arise not from linguistic deficits, but from motor planning failures in Broca’s area and the supplementary motor area, where 5HT2A signaling modulates corticobulbar projections to cranial nerves that control articulation, including hypoglossal output for tongue control.
When the motor plan cannot be reliably executed, speech fails even when comprehension is intact. That was Jacob. That is many non speaking autistic individuals who type with advanced language and insight, like we have seen in the Speller’s Documentary by JB Hadley and Dana Johnson. That is why autism can look like low cognition while hiding complex cognition behind an output blockade.
If you accept the concept of microbiome driven psychedelic apraxia, then many classic autism traits become secondary effects of disrupted motor planning, disrupted gating, and disrupted salience assignment.
In the next section, we widen the lens to the symptom clusters families live with every day: OCD loops, aggression, sleep disruption, and seizures, and we explore how the same chemistry can plausibly unify them.
Section 8: OCD Loops, Aggression, Sleep disruption, and Seizures, One Circuit Logic
Parents who have a severely autistic child often describe a pattern that feels like a storm system: periods of relative stability followed by cycles of insomnia, obsessive rigidity, escalating agitation, aggression, self injury, and sometimes seizures.
A model that only labels autism as “social deficits and repetitive behaviors” does not do justice to this reality. The lived phenotype is a neurophysiological volatility syndrome.
A chronic endogenous psychedelic state can plausibly generate this volatility through several converging mechanisms:
First, sensory flooding drives chronic stress arousal. When gating fails, the nervous system lives closer to fight or flight.
Second, 5HT2A driven cortical excitation increases network noise and destabilizes inhibitory control. That can manifest as impulsivity, irritability, and aggression, especially in individuals already vulnerable through inflammation, mitochondrial stress, or toxin burden.
Third, the cortico striato thalamo cortical loops that govern motor habits and cognitive flexibility can become stuck. That looks like OCD, perseveration, and rigid loops. In many individuals, the repetitive behaviors may function as self generated rhythmic regulation in a world that feels unpredictably amplified.
Fourth, sleep becomes a direct biochemical casualty. Tryptophan is substrate. When the microbiome hijacks tryptophan into tryptamine pathways, serotonin and melatonin dynamics can be altered. Sleep fragmentation, early waking, circadian instability, and night agitation become predictable outputs of upstream chemistry.
Fifth, seizure threshold can drop. Cortical hyperexcitability, altered thalamocortical oscillations, neuroinflammation, and disrupted calcium signaling can converge on a nervous system that is more likely to seize. Clinically, many families report that when gut driven inflammation and dysbiosis improve, seizure frequency and severity often improve as well. We have observed this repeatedly among families implementing a structured Gut Balancing program (more info: https://www.autismisbiomedical.com/gutbalancing).
In the next section, we connect this to objective brain findings, MRI and EEG patterns, because autism has repeatedly shown early brain overgrowth and connectivity abnormalities that fit the concept of chronic network destabilization.
Section 9: MRI and EEG patterns that fit an Altered Consciousness Model
Young autistic brains often show early overgrowth patterns on MRI. Many studies describe larger total brain volume in early childhood in subsets of autism, and abnormal developmental trajectories thereafter. Functional imaging frequently shows atypical connectivity patterns, sometimes local hyperconnectivity, sometimes long range underconnectivity, often an imbalance that depends on age and phenotype.
The Brain Overgrowth Phenomenon
One of the most consistent biological findings in autism is early brain overgrowth. Approximately 15-20% of children with autism exhibit macrocephaly (abnormally large head size), but MRI studies reveal that brain volume overgrowth is far more common, affecting a majority of toddlers with autism before normalizing or degenerating later in childhood. I do not believe inflammation of the brain or any toxin could make a brain larger. Please correct me, I could be wrong.
- Volumetric Surge: Between the ages of 2 and 4 years, autistic brains can be significantly larger—sometimes exceeding norms by 10% (equivalent to 200-400 grams of excess tissue). This overgrowth is most pronounced in the frontal and temporal lobes (areas critical for language, social cognition, and motor planning) and the cerebellum.
- Mechanism: Synaptic Pruning Failure: Normal development requires “synaptic pruning”—the elimination of weak or redundant connections to sharpen neural circuits. 5HT2A signaling is a powerful driver of neuroplasticity and dendritic growth (via BDNF and mTOR pathways). If a child’s brain is chronically bathed in endogenous 5HT2A agonists (like DMT/bufotenin) during critical developmental windows, the signal for “growth and connection” may overwhelm the signal for “pruning and refinement.”
This results in a “noisy,” hyper-connected local network structure that is biologically expensive and functionally inefficient. Exactly the pattern observed in ASD neuroimaging.
Now consider what psychedelics do in imaging studies. Psychedelics are associated with increased network flexibility, altered effective connectivity, disrupted default mode network integrity, altered thalamocortical coupling, and changes in measures that correlate with “entropy” or signal diversity.
The conceptual overlap matters. If a brain is chronically exposed to psychedelic like signaling through 5HT2A and sigma 1 pathways, you would predict:
- Altered oscillatory rhythms, especially reduced alpha suppression and abnormal sensory gating dynamics
- Altered thalamocortical connectivity
- Altered balance between top down prediction and bottom up sensory influx
- Periods of hyperconnectivity in developing networks, potentially linked to altered synaptic pruning and plasticity signaling
- A behavioral phenotype that looks like sensory amplification, motor planning disruption, and unstable regulation
This does not require that autism equals a psychedelic trip. It requires a subtler claim: a chronic microdose like neurochemical environment can distort network development and real time gating, producing a phenotype that mimics, in slow motion, the same circuit disruptions seen in acute psychedelic states.
In the next section, we stop describing and start predicting, because the strength of a hypothesis is not its poetry, it is its ability to forecast who will respond, what patterns will be seen, and what interventions will matter most.
Section 10: Predictions, the Responder Profile, and Why this is Likely not a Niche Subgroup
A mechanistic hypothesis should produce a clinical fingerprint. This one does.
The predicted responder profile includes many of the families who are already deep into biomedical work, because they are the ones who have noticed the gut brain connection relentlessly. The pattern includes:
- Significant GI dysfunction, constipation, diarrhea, abdominal pain, reflux, food reactivity
- Sleep disruption, early waking, night agitation, circadian instability
- Sensory overwhelm, noise intolerance, tactile defensiveness, startle reactivity
- Aggression, irritability, self injury, emotional volatility that cycles
- OCD loops, rigid perseveration, intense insistence on sameness
- Marked dyspraxia, poor coordination, speech apraxia, non speaking status despite strong comprehension
- Seizures or subclinical epileptiform patterns, or obvious seizure threshold fragility
- Evidence of dysbiosis on stool analysis, often including depletion of protective Bifidobacterium species and depleted Lactobacillus species, alongside expansion of taxa with strong tryptophan decarboxylase capacity that convert tryptophan into tryptamine
A common objection would be that this might apply to only a small subgroup. The clinical reality argues otherwise. Autism, in the real world, is saturated with gut dysfunction, sleep disruption, sensory gating problems, and motor planning impairment. Those are not fringe features. They are core features in the majority of moderate to severe cases, and present in many higher functioning cases in more subtle ways.
Therefore the more accurate framing is: endogenous psychedelic modulation likely oscillates in degree across the spectrum, and across time, and across triggers. Most individuals with autism may not be in a constant maximal state. They may fluctuate with diet changes, infections, antibiotics, stress, constipation severity, motility changes, fungal shifts, toxin exposures, and inflammation. That fluctuation alone would explain why many parents describe waxing and waning cognition, waxing and waning eye contact, waxing and waning sleep, and sudden behavioral storms that appear and disappear.
In the next section, we move from prediction to action, because this hypothesis does not exist as an intellectual exercise. It exists because a structured intervention has already produced meaningful changes in hundreds of families, and those outcomes demand a mechanistic explanation.
Section 11: Gut Balancing, what Alex built, what he Saw, and Why it Matters
Gut Balancing is a six week personalized intervention program developed by my partner, Alex Zaharakis. Alex has analyzed over 2000 16S stool samples over the years and built a system that translates microbial patterns into targeted, structured suggestions using botanicals, herbals, prebiotics, probiotics, and nutraceutical supports. The goal is not to chase a mythical perfect microbiome. The goal is to reduce the specific overgrowths driving inflammatory and neuroactive output, and to rebuild protective function, especially through restoration of barrier support and keystone organisms.
Hundreds of families have implemented this program and reported meaningful improvements. This includes families dealing with severe phenotypes, including seizure prone children, children with profound speech impairment, and children with relentless insomnia and aggression cycles.
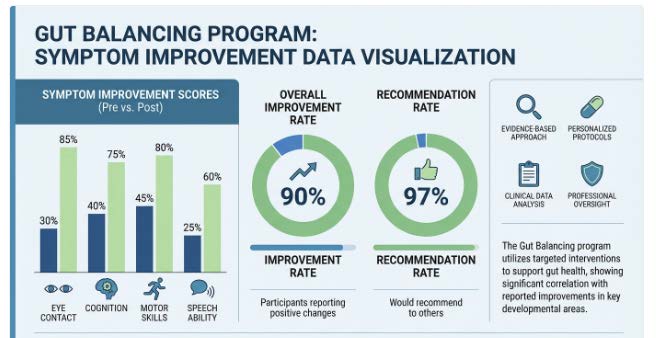
We analyzed outcomes in a retrospective dataset of 219 participants, ages 2 to 29, who completed at least one six week round. In the primary survey cohort, over 90 percent of caregivers reported overall improvement, 97 percent would recommend Gut Balancing to other parents, and 96 percent expressed interest in doing an additional round. The symptom areas with particularly strong improvement to worsening ratios included eye contact, cognition, joint attention, motor coordination, and verbal speech ability.
This is not a claim that one program “treats autism.” It is a claim that the gut brain axis is not theoretical. It is modifiable, and when it is modulated intelligently, core autism traits can move. That reality is already forcing a new scientific question: what specific biochemical outputs are being reduced when families improve, and what does that reveal about the causal chemistry of the autism phenotype?
This is where upstream intervention logic becomes unavoidable. Jacob’s insights catalyzed a wave of families implementing targeted detoxification strategies, including sodium thiosulfate among a broader arsenal of precision supports. The mechanistic elegance here is that supporting detoxification capacity, restoring GI motility, and rebalancing the microbiome ecosystem converge on a single biological outcome: reduced aberrant molecular flux across a compromised intestinal barrier. When barrier integrity is restored, the cascade reverses: inflammatory signaling attenuates, hepatic biotransformation capacity normalizes, mitochondrial energetics stabilize, and the nervous system regains access to homeostatic operating parameters. The phenotype softens not because we “treated autism,” but because we interrupted the biochemical drivers generating the altered state.
In the next section, the narrative closes the loop: why we are now formalizing this into a research program with high level collaborators, and why the stakes extend beyond autism into a new biology of consciousness and neuropsychiatric illness.
Section 12: Why we are Taking this to the Laboratory Now
This article exists for a specific reason. It is not written to provocate the status quo of the conventional medicine model. It is written because the clinical signal has become too coherent to ignore, and the hypothesis is now testable.
We are planning to study these concepts with Dr. Michael Angelo at Stanford, Steven Barker, PhD at LSU, Dr. Diane Hennacy, and David Olson PhD at UC Davis. The purpose is not to turn autism into a metaphysical story. The purpose is to map a biochemical reality: whether dysbiotic microbiomes in autism are producing an excess indole tryptamine ecosystem that can chronically alter brain function.
A secondary purpose is equally urgent: to assess whether individuals with severe autism may have far higher intelligence and comprehension than commonly assumed, and whether what looks like “cognitive impairment” is often a motor output disorder, a biological apraxia driven by disrupted motor planning circuits. If the interface is broken while the mind is intact, then the ethical implications for education, care, and civil rights are enormous.
This also matters for policy. The current national approach to autism is dominated by services, behavior management, and limited pharmacology. The FDA has two commonly used medications for autism irritability, and both prominently block 5HT2A signaling. That should not be the end of the story. It should be the beginning of a deeper investigation into why that receptor is relevant in the first place, and whether the upstream driver is modifiable at the gut level, with a precision systems biology approach rather than lifelong sedation.
For anyone interested in getting involved and/or funding support, please email me at cbogner@drbognerhealth.com.
This Endogenous Psychedelic Hypothesis, and the concept of microbiome driven psychedelic apraxia, offers a unifying path: gut dysbiosis drives tryptamine production, barrier dysfunction increases absorption, enzymology converts substrates into DMT and bufotenin like compounds, genetics can reduce clearance, chronic receptor activation reshapes sensory gating, motor planning, sleep, emotional regulation, and seizure threshold, and the outward phenotype converges on what we call autism.
If this is correct, the future of autism research and treatment changes. Autism stops being a static label and becomes a dynamic biochemical state with measurable drivers, predictable fluctuations, and mechanistically rational interventions.
Resources:
Rumble Channel
https://rumble.com/user/ResearchedElements
Dr. Bogner on X
Dr. Bogner on Facebook
https://www.facebook.com/DrBogner
Researched Elements
https://www.researchedelements.com
Autism is Biomedical Foundation
https://www.autismisbiomedical.com
Testimonials
https://www.researchedelements.com/testimonials
Gut Balancing
https://www.autismisbiomedical.com/gutbalancing
Parent Interviews
https://www.autismisbiomedical.com/hearfromparents
These links provide educational resources, research updates, and firsthand accounts from families who have implemented similar interventions.
You can review experiences and outcomes from other parents through these links:
Parent emails: link
Video testimonials: link
Additional testimonials: link
Program outcomes: link
Program details: Gut Balancing
Additional resources include parent interviews (YouTube 1, YouTube 2, YouTube 3), real-time survey statistics (ASD group), Biomesight practitioner reviews (link), and additional educational material (Rumble).